生命科学を支える 細胞生物学
News / Topics
新着お知らせ
-
CSF論文発表のお知らせ
Opto-p53: A Light-Controllable Activation of p53 Signaling Pathway
この論文では、がん抑制タンパク質p53の活性を光制御するツールを開発し、細胞周期停止や細胞死を誘導できることを示しました。
-
CSF論文発表のお知らせ
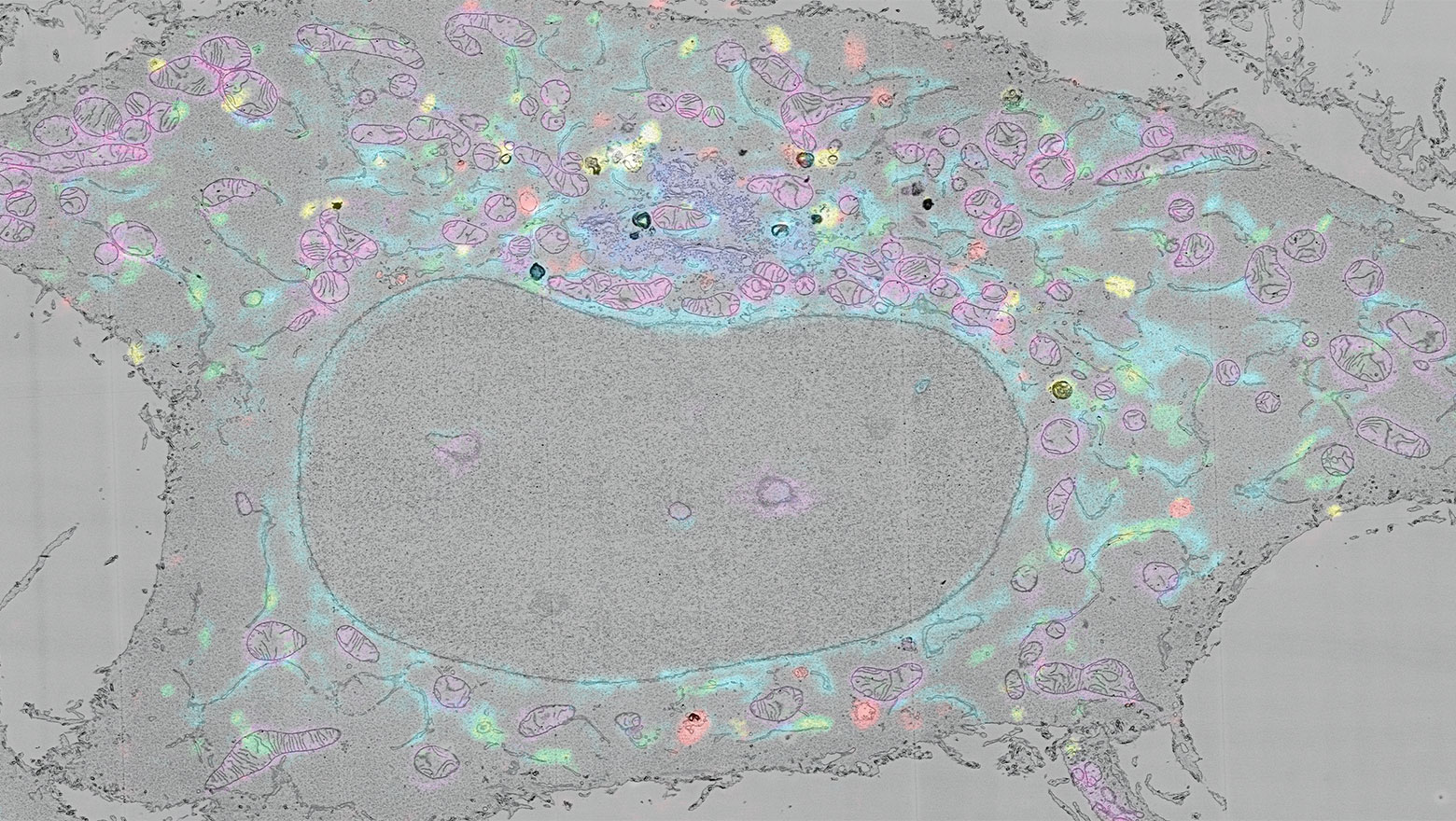
Cell biological insights into human STING variants
この論文は、自然免疫分子STINGのバリアントの機能と疾患との関連についてまとめた総説です。
-
CSF論文発表のお知らせ
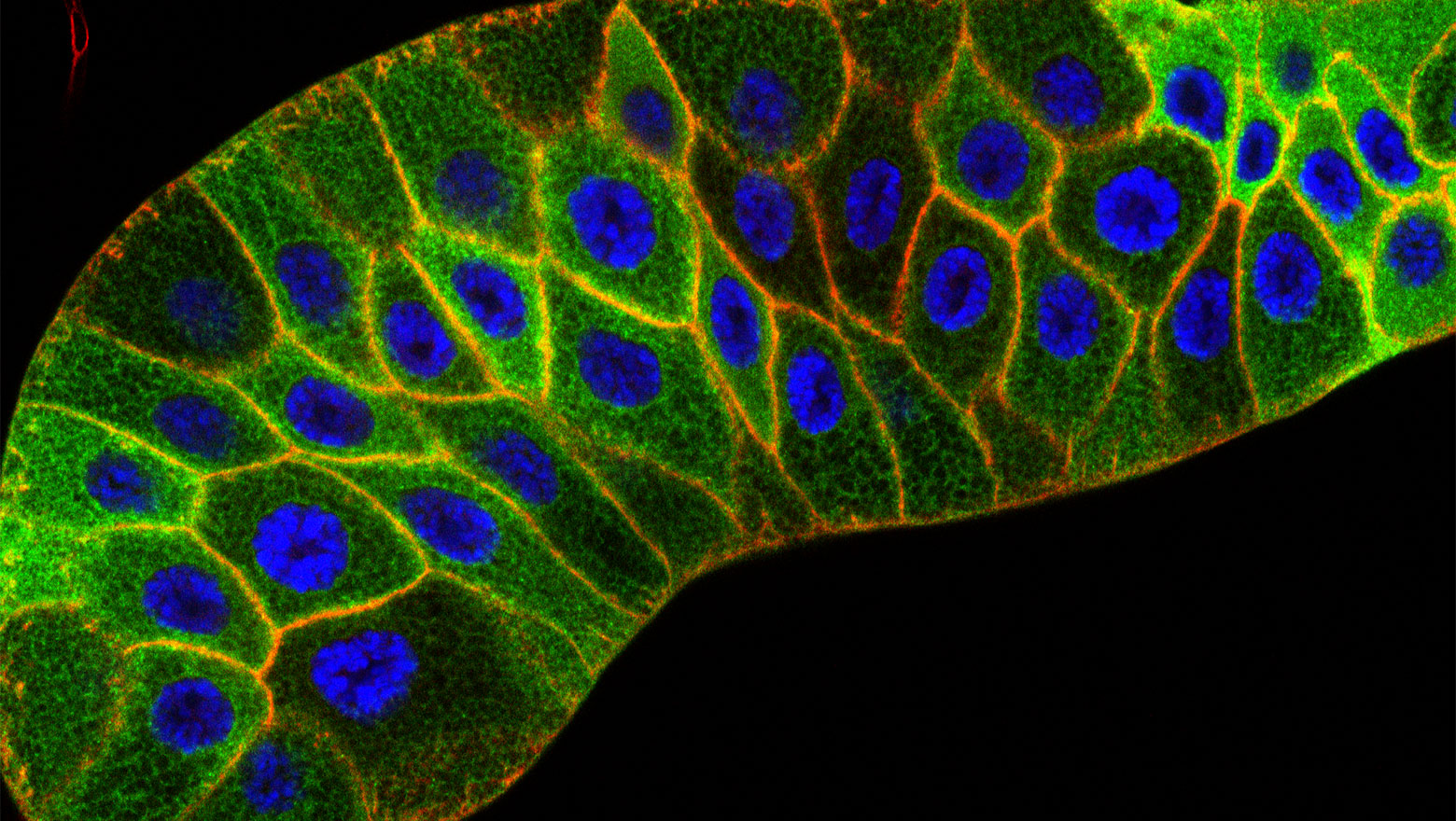
BiP, GRP94, Calreticulin and Calnexin Contribute to Development of the Notochord in Medaka Fish
この論文は、小胞体局在性の分子シャペロンであるBiP, GRP94, calreticulinとcalnexinがメダカの脊索発達に寄与することを示した。
-
CSF論文発表のお知らせ
A quantitative method to monitor STING degradation with dual-luciferase reporters
この論文では、自然免疫分子STINGの分解を定量するハイスループットアッセイ系を構築しました。
-
CSF論文発表のお知らせ
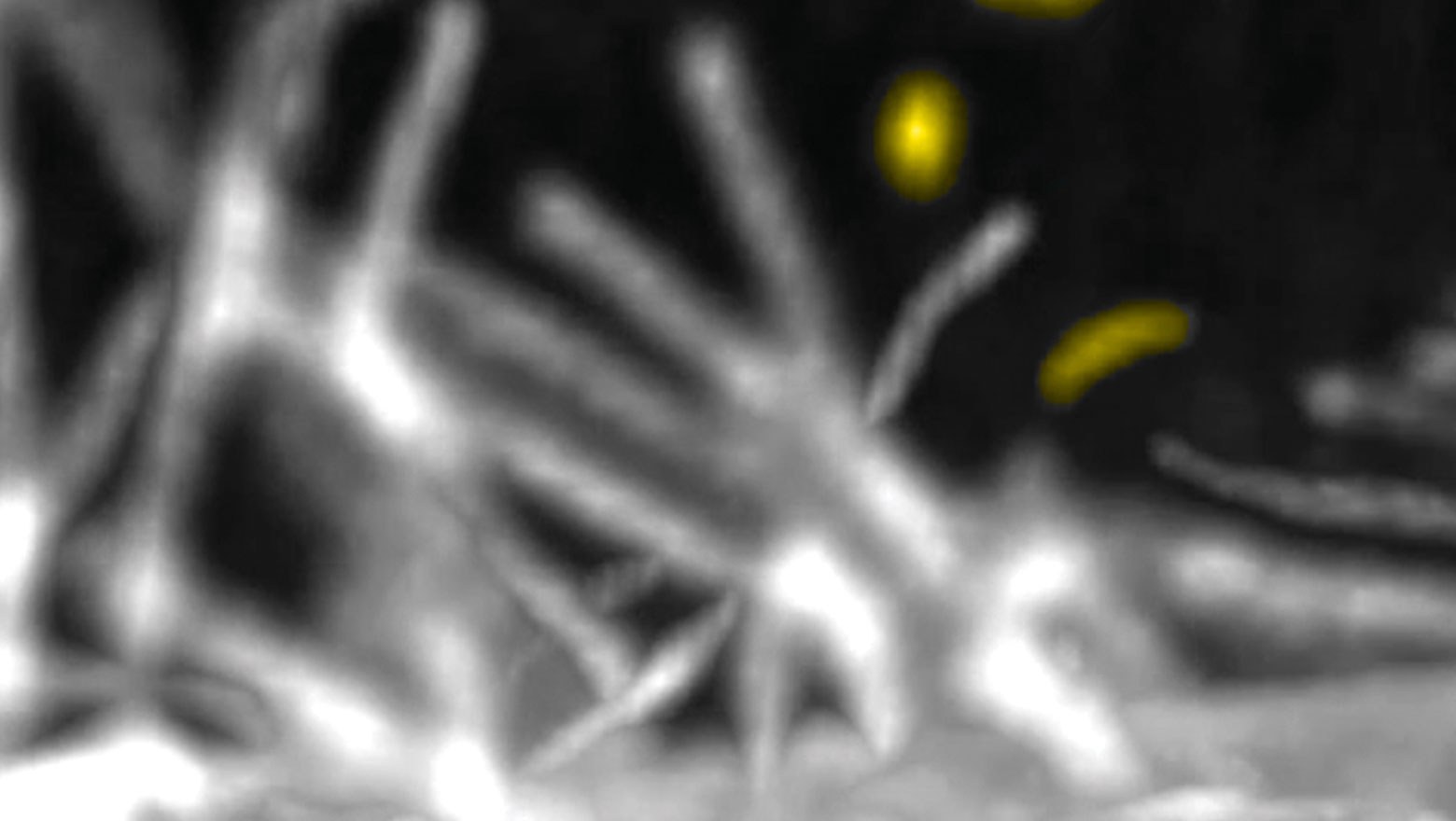
Chemokine induces phase transition from non-directional to directional migration during angiogenesis
この論文では、脳の毛細血管網形成において新生血管が標的血管へ向かって移動する機構を報告しています。
Journal
会報「細胞生物」
Events
学術大会・イベント
About
当学会について